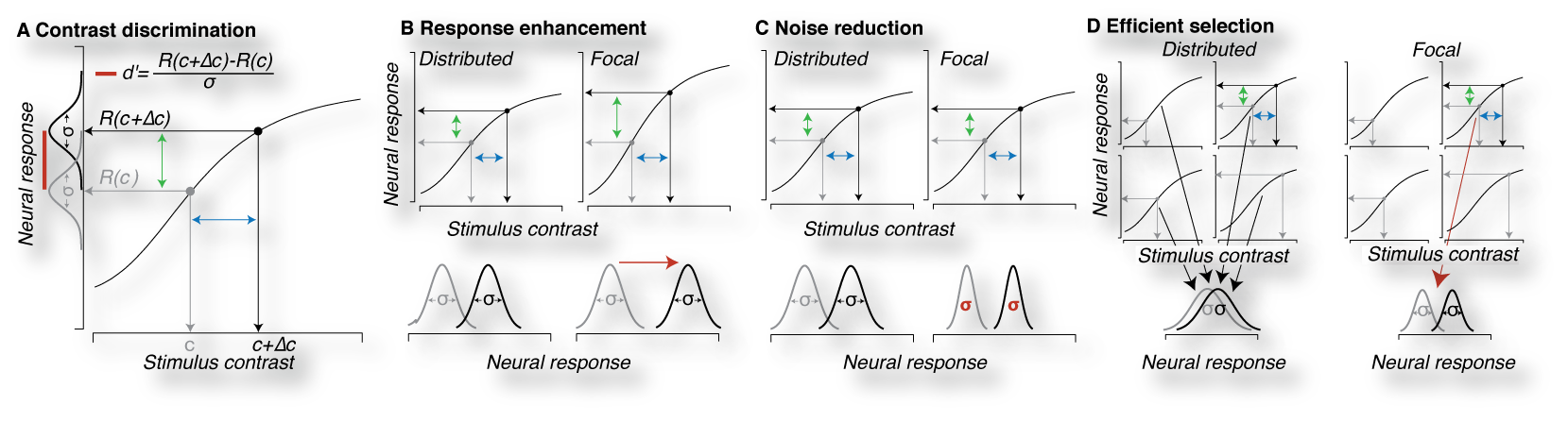
 Our approach is to connect measurements of human cortical brain activity to behavior and perception through the use of linking models (Gardner, 2015). Linking models are formal, computational descriptions of hypothesized mechanisms by which measured brain activity is transformed into perception or behavior. Linking models instantiate specific hypotheses about cortical mechanisms that can be empirically tested by seeing whether they can account for observed physiology and behavior. While cognitive neuroscience has produced numerous descriptions of brain processes that are potentially “involved in” or “associated with” cognitive functions, it is critical to transform such descriptions into computational linking models of brain mechanisms which make explicit, falsifiable predictions that can be generalized and tested in novel circumstances. Such linking models can rigorously support links between brain and behavior, as well as, on occasion, lead to unexpected conclusions. Moreover, linking models form the foundation for a cumulative, replicable and expanding body of cognitive neuroscience knowledge because they can be repeatably tested with new data, amended as they fall short of capturing novel phenomenon and generalized so that they can make useful predictions. As such models gain predictive validity, they will have broad societal impacts by being the basis for how engineers evaluate how system design decision can complement and enhance human cognition, or how doctors diagnose, or educators intervene to improve cognitive function. Our approach is to connect measurements of human cortical brain activity to behavior and perception through the use of linking models (Gardner, 2015). Linking models are formal, computational descriptions of hypothesized mechanisms by which measured brain activity is transformed into perception or behavior. Linking models instantiate specific hypotheses about cortical mechanisms that can be empirically tested by seeing whether they can account for observed physiology and behavior. While cognitive neuroscience has produced numerous descriptions of brain processes that are potentially “involved in” or “associated with” cognitive functions, it is critical to transform such descriptions into computational linking models of brain mechanisms which make explicit, falsifiable predictions that can be generalized and tested in novel circumstances. Such linking models can rigorously support links between brain and behavior, as well as, on occasion, lead to unexpected conclusions. Moreover, linking models form the foundation for a cumulative, replicable and expanding body of cognitive neuroscience knowledge because they can be repeatably tested with new data, amended as they fall short of capturing novel phenomenon and generalized so that they can make useful predictions. As such models gain predictive validity, they will have broad societal impacts by being the basis for how engineers evaluate how system design decision can complement and enhance human cognition, or how doctors diagnose, or educators intervene to improve cognitive function. |
 Even our most ordinary of visual experiences requires extraordinary computational feats of the human brain. Everyday visual scenes are a riot of clutter and confusion that require our perceptual systems to selectively attend to what is relevant and to infer from uncertain sensory information the state of the world. The overarching goal of our research is to understand the human brain mechanisms that give rise to such extraordinary cognitive feats as selective attention and sensory inference.
Even our most ordinary of visual experiences requires extraordinary computational feats of the human brain. Everyday visual scenes are a riot of clutter and confusion that require our perceptual systems to selectively attend to what is relevant and to infer from uncertain sensory information the state of the world. The overarching goal of our research is to understand the human brain mechanisms that give rise to such extraordinary cognitive feats as selective attention and sensory inference.  Our approach is to connect measurements of human cortical brain activity to behavior and perception through the use of linking models (Gardner, 2015). Linking models are formal, computational descriptions of hypothesized mechanisms by which measured brain activity is transformed into perception or behavior. Linking models instantiate specific hypotheses about cortical mechanisms that can be empirically tested by seeing whether they can account for observed physiology and behavior. While cognitive neuroscience has produced numerous descriptions of brain processes that are potentially “involved in” or “associated with” cognitive functions, it is critical to transform such descriptions into computational linking models of brain mechanisms which make explicit, falsifiable predictions that can be generalized and tested in novel circumstances. Such linking models can rigorously support links between brain and behavior, as well as, on occasion, lead to unexpected conclusions. Moreover, linking models form the foundation for a cumulative, replicable and expanding body of cognitive neuroscience knowledge because they can be repeatably tested with new data, amended as they fall short of capturing novel phenomenon and generalized so that they can make useful predictions. As such models gain predictive validity, they will have broad societal impacts by being the basis for how engineers evaluate how system design decision can complement and enhance human cognition, or how doctors diagnose, or educators intervene to improve cognitive function.
Our approach is to connect measurements of human cortical brain activity to behavior and perception through the use of linking models (Gardner, 2015). Linking models are formal, computational descriptions of hypothesized mechanisms by which measured brain activity is transformed into perception or behavior. Linking models instantiate specific hypotheses about cortical mechanisms that can be empirically tested by seeing whether they can account for observed physiology and behavior. While cognitive neuroscience has produced numerous descriptions of brain processes that are potentially “involved in” or “associated with” cognitive functions, it is critical to transform such descriptions into computational linking models of brain mechanisms which make explicit, falsifiable predictions that can be generalized and tested in novel circumstances. Such linking models can rigorously support links between brain and behavior, as well as, on occasion, lead to unexpected conclusions. Moreover, linking models form the foundation for a cumulative, replicable and expanding body of cognitive neuroscience knowledge because they can be repeatably tested with new data, amended as they fall short of capturing novel phenomenon and generalized so that they can make useful predictions. As such models gain predictive validity, they will have broad societal impacts by being the basis for how engineers evaluate how system design decision can complement and enhance human cognition, or how doctors diagnose, or educators intervene to improve cognitive function.